Structural Diversity
Trees, herbs, vines, ferns — every growth form fills a different role. When your garden mixes shapes, it captures more light, shelters more predators, buffers temperature, and stays productive through bad years.
In plain terms: a garden with five shapes of plant can outperform a garden of fifty plants that all look the same.

Low diversity
All herbs, all the same height. One light zone, one root depth, bare soil between plants. If a drought hits, everything tends to fail together.
High diversity
Tree canopy, shrub middle, herb floor, vine climbing, fern in shade. Five layers capturing light, five root depths tapping water, five ways to survive a bad year.
The Core Finding
In one of ecology's most replicated results, David Tilman and colleagues showed in 1997 that the number of different plant shapes — trees, grasses, herbs, legumes — matters more than the total species count. When they tested six measures of how well the ecosystem worked (plant growth, nutrient use, soil health, and more), the mix of growth forms was significant for all six. The number of species? Significant for none.
Peter Reich extended this in 2004 with a cleverly designed experiment that separated the effects of growth form and species variety. Both independently boosted plant growth — growth forms by 28%, species by 40%. The key finding: these are separate, additive effects. Growth form diversity and species diversity each drive complementarity — plants helping each other — through different mechanisms. You get the most out of a garden when you mix both shapes and species.
What this means for your garden: A guild of 5 plants from 4 different growth forms (tree, shrub, herb, grass) will likely outperform 5 plants that are all herbs — even if the herb-only mix has more species. Growth form diversity is the structural foundation.
Tilman et al. 1997; Reich et al. 2004
A — The Productivity Engine
Gardens and forests with a mix of heights and shapes capture more light — and more light means more growth. This isn't a small effect. Worldwide, the physical structure of the canopy predicts total plant growth better than the number of species or the total amount of leaf.
83%
of productivity variance
Explained by canopy bumpiness
vs 31% for total leaf cover, 30% for species count
88%
more productive
Complex vs simple stands
Ray et al. 2023, MyDiv experiment
78%
of ecoregions
Show the pattern worldwide
Liu et al. 2024, 496 forest ecoregions
Gough et al. 2019; Ray et al. 2023
Different shapes grab light at different heights: a tree captures the top, a shrub grabs the middle, a herb uses what filters through, and a vine climbs to find its own patch. When grown alongside other shapes, trees grow 38% larger crowns than when planted alone (Jucker et al. 2015). They literally reshape themselves to fill the gaps — nature's version of efficient packing.
The bumpiness of the canopy — how much the treetops varied in height — explained 83% of total plant growth across 10 US forest sites. That's nearly three times more than total leaf cover (31%) or species count (30%). When researchers tested all three factors together, only the bumpiness mattered. (Gough et al. 2019)
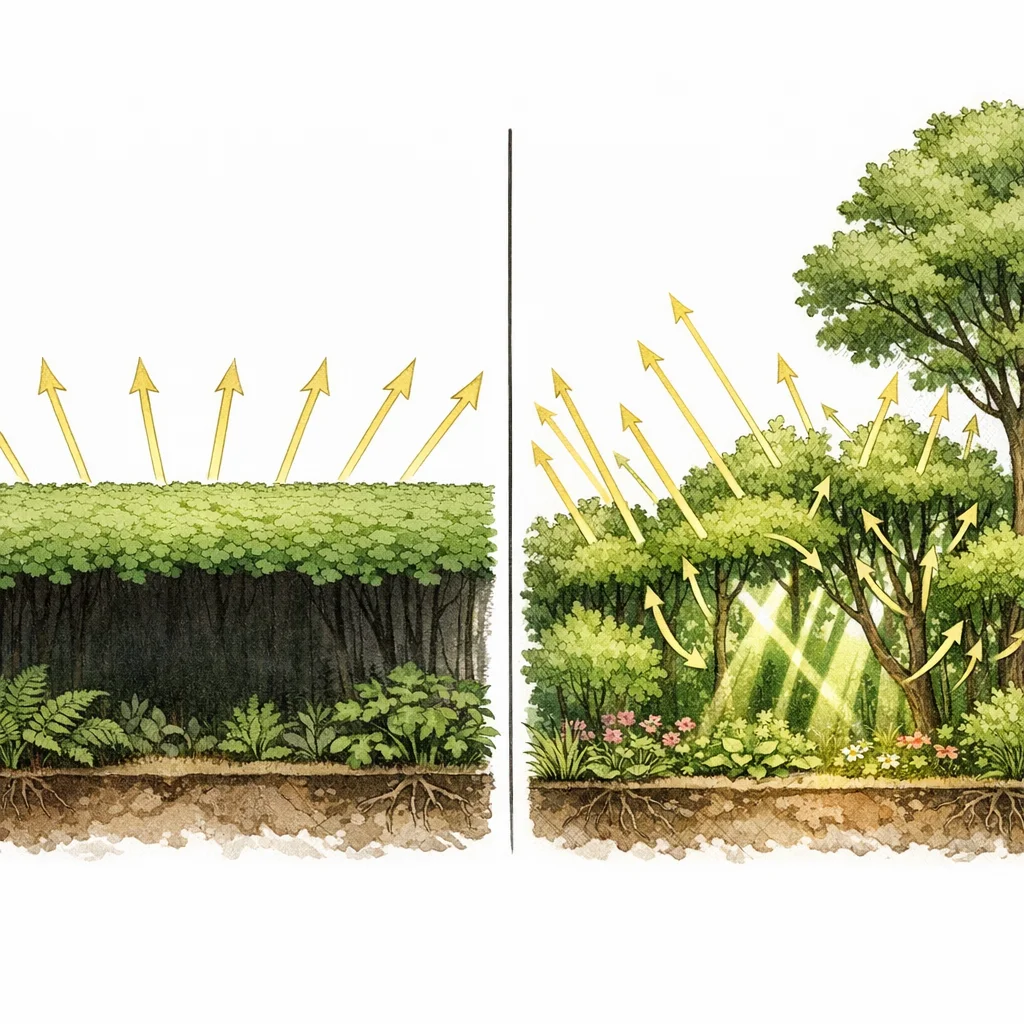
Liu et al. (2024) used satellite lidar measurements of forest structure across 496 ecoregions worldwide and confirmed: structurally complex forests are more productive in 78% of them. The pattern holds from cold northern forests to the tropics — this isn't a local quirk. The effect was even stronger in managed forests, where structural complexity tends to start lower — suggesting the biggest gains come from adding variety where it's currently lacking.
B — The Pest Control Layer
A garden with structural complexity is a garden with hiding places — for the insects, spiders, and birds that eat your pests. The more physical variety in your planting, the more types of beneficial predators find a home.
Large
boost to beneficial insects
Consistent across 43 studies
Langellotto & Denno 2004
120×
more pests
In simple vs diverse gardens
Shrewsbury & Raupp 2006
Langellotto & Denno 2004; Shrewsbury & Raupp 2006
The single strongest feature for attracting pest-eating predators? Leaf litter and mulch on the ground. Hunting spiders need it for refuge from cannibalism and intraguild predation. Web-building spiders need physical anchor points between stems and branches to build webs. A garden with multiple layers of planting provides both (Langellotto & Denno 2004).
Garden-scale proof. Pest numbers on azaleas were 120 times higher in simple, tidy urban landscapes than in complex ones with mixed planting. And the rate at which predators ate those pests was 3.3× higher where structural complexity remained. This is one of the few studies conducted at actual garden scale. (Shrewsbury & Raupp 2006)

Barnes et al. (2020) modelled 487 who-eats-whom networks and found that 16-species mixtures supported 162% more predator activity than single-species plantings — and herbivore feeding rates dropped by 44%. The reason is intuitive: more plant shapes means more types of hiding places, hunting grounds, and web-building sites for different predators.
In a large Chinese forest experiment, Schuldt et al. (2019) found that structural diversity was the dominant factor explaining the species richness of predators and parasitoid wasps — more than leaf functional traits. In forests, the architecture of the planting mattered more than the traits of individual tree species (though in grasslands, functional diversity played a larger role).
C — The Temperature Shield
Multiple vegetation layers don't just capture light — they buffer temperature. On hot days, the canopy shades the ground; on cold nights, the layers trap warmth.
4.4°C
cooler in summer
24-species vs monocultures
+ 1.1°C warmer in winter
−1.94°C
per vegetation layer
Temperature reduction per layer
Terschanski et al. 2024, 17 sites
Schnabel et al. 2025
Schnabel et al. (2025) ran a 6-year experiment across 64 forest plots in China. Mixtures with 24 species were 4.4°C cooler at summer maximum and 1.1°C warmer at winter minimum compared to single-species plots. When they tested why, two structural features explained the entire effect: how dense the leaf canopy was, and how varied the heights were. Once those two structural features were included, species diversity added nothing extra — shape IS the mechanism.
Cooler summers AND warmer winters. Structural diversity creates a microclimate buffer that works in both directions — protection from heat stress during heatwaves and from frost during cold snaps. This is especially valuable as climate extremes become more frequent.
Separately, Terschanski et al. (2024) measured 17 sites across an Afromontane land-use gradient and found the mix of vegetation heights mattered more than the density of the canopy overhead. Each additional vegetation layer — ground cover, shrubs, understory trees, canopy — reduced the gap between daytime high and nighttime low by nearly 2°C. It's not about how much leaf there is — it's about how many layers it's spread across.
D — The Insurance Policy
Even if nothing goes wrong this year, structural diversity is insurance for when something does.
Different growth forms respond differently to stress. Drought can kill shallow-rooted herbs while deep-rooted trees may be buffered. A late frost destroys tender new growth but bulbs and tubers dormant underground don't notice. Wind is more likely to flatten a uniform stand but can dissipate through mixed heights. Because different shapes fail at different times, the garden as a whole keeps going — ecologists call this the "insurance effect."
Diversity buffers the damage. Diverse communities show significantly less variability in productivity during stress events than low-diversity ones. The insurance effect is one of the best-supported findings in biodiversity research, backed by strong evidence from both theory and field experiments. (Loreau et al. 2021)
Yachi & Loreau (1999) showed mathematically that the buffering benefit of additional species saturates — each new species helps, but the biggest gains come from the first few. Because different growth forms tend to be functionally different — different rooting depths, different tolerances, different growth speeds — mixing shapes is an effective way to build this insurance. Our 12-category system captures the key shape differences (tall tree vs low herb vs underground bulb vs climbing vine) that determine how a plant responds to heat, frost, drought, or wind.
Theory identifies three mechanisms that stabilise diverse ecosystems (Loreau & de Mazancourt 2013), and mixing growth forms activates all three. First, different shapes have different tolerances — they don't all fail in the same heatwave or drought. Second, fast and slow growers complement each other: an annual herb bounces back in months while a tree takes decades but provides stable structure year-round. Third, because each shape taps different resources — different soil depths, different light levels — they don't compete as directly with each other.
How We Measure It
We use Shannon's diversity index — a mathematical formula that rewards both the number of growth forms and how evenly they're distributed. A guild with 3 trees, 3 herbs, and 3 ferns scores higher than one with 8 trees and 1 herb, even though both have the same total plants.
Classify every plant
We assign each species one of 12 growth forms using the World Checklist of Vascular Plants (WCVP v15). WCVP classifies plants using a modified Raunkiaer system that pairs each growth form (structural habit) with a Raunkiaer life form (position of perennating buds) — the same two axes used by Taylor et al. (2023) across 295,000+ species. We extend WCVP’s base categories with three family-level assignments: palm, graminoid, and fern.
| Growth form | Raunkiaer life form | WCVP definition |
|---|---|---|
| Tree | Phanerophyte | Woody stems persisting for several years; buds normally above 3 m |
| Shrub | Nanophanerophyte | Woody stems persisting for several years; buds above soil level but normally below 3 m |
| Subshrub | Chamaephyte | Herbaceous and/or woody stems persisting for several years; buds on or just above soil level, never above 50 cm |
| Herb | Hemicryptophyte | Herbaceous stems, often dying back after growing season; buds just on or below soil level |
| Herb (annual) | Therophyte | Survives unfavourable seasons as seeds; completes life cycle during favourable season |
| Geophyte | Geophyte | Survives unfavourable seasons via underground storage organs (rhizome, bulb, tuber, or root bud) |
| Vine | modifier | Climbing — applied as a modifier across life forms (e.g. cl. phan., cl. hemicr.) |
| Epiphyte | modifier | Growing on trees or rocks, without extracting nutrition from the host. Final authority: EpiList (Zotz et al. 2021, 24,279 species) |
| Succulent | modifier | WCVP base noun or modifier on herb/shrub/subshrub; gap-filled from TRY database |
| Bamboo | Phanerophyte | Listed under “Others” in WCVP glossary; tagged directly in lifeform codes |
| Palm | Phanerophyte | Family Arecaceae — assigned by family membership |
| Graminoid | — | Poaceae, Cyperaceae, Juncaceae, Restionaceae, Typhaceae — assigned by family (APG IV) |
| Fern | — | 53 pteridophyte families incl. lycophytes — assigned by family (PPG I 2016 + worldplants.de v26.02) |
Definitions from WCVP/POWO lifeform glossary (Raunkiaer 1934, modified per De Langhe et al. 1983). Epiphyte authority: EpiList (Zotz et al. 2021). Succulent gap-fill: TRY database. Three non-structural categories excluded from scoring: aquatic (Taylor 2023 excluded hydrophytes), parasite, mycotroph. Family lists: ferns — worldplants.de, Pteridophyte Phylogeny Group I (2016); graminoids — Angiosperm Phylogeny Group IV (2016).
Count the mix
For your guild, we tally how many plants fall in each category and calculate the proportions. A guild of 10 plants — 4 trees, 3 herbs, 2 ferns, and 1 vine — has four forms with proportions 0.4, 0.3, 0.2, and 0.1.
Score the diversity
The formula turns those proportions into a single score. A higher score means more forms present and more evenly spread across them. We convert this to a 0–100 scale, where 100 would be a perfectly even mix of all 12 forms.
What This Metric Doesn't Capture
The Research Behind It
This is not one study. It is a converging body of 22 peer-reviewed papers — including large-scale reviews, field experiments across multiple continents, and global satellite analysis — spanning three decades:
"Canopy rugosity [bumpiness] explained 83% of net primary productivity [total plant growth] — more than species richness, leaf area index, or any trait we measured."
"Structural complexity increased natural enemy abundance with a large, consistent effect across 43 studies, driven primarily by ground-level habitat features."
"Twenty-four-species mixtures were 4.4°C cooler in summer and 1.1°C warmer in winter — fully mediated by canopy density and structural complexity."