Soil Biodiversity
Beneath every thriving guild lies two invisible communities working in parallel — fungi expanding every root's reach into the soil, and bacteria fixing the very air into food for your plants.
Fungi
Mycorrhizal partnerships and fungal disease control
Bacteria
Nitrogen-fixing Rhizobium and free-living fixers
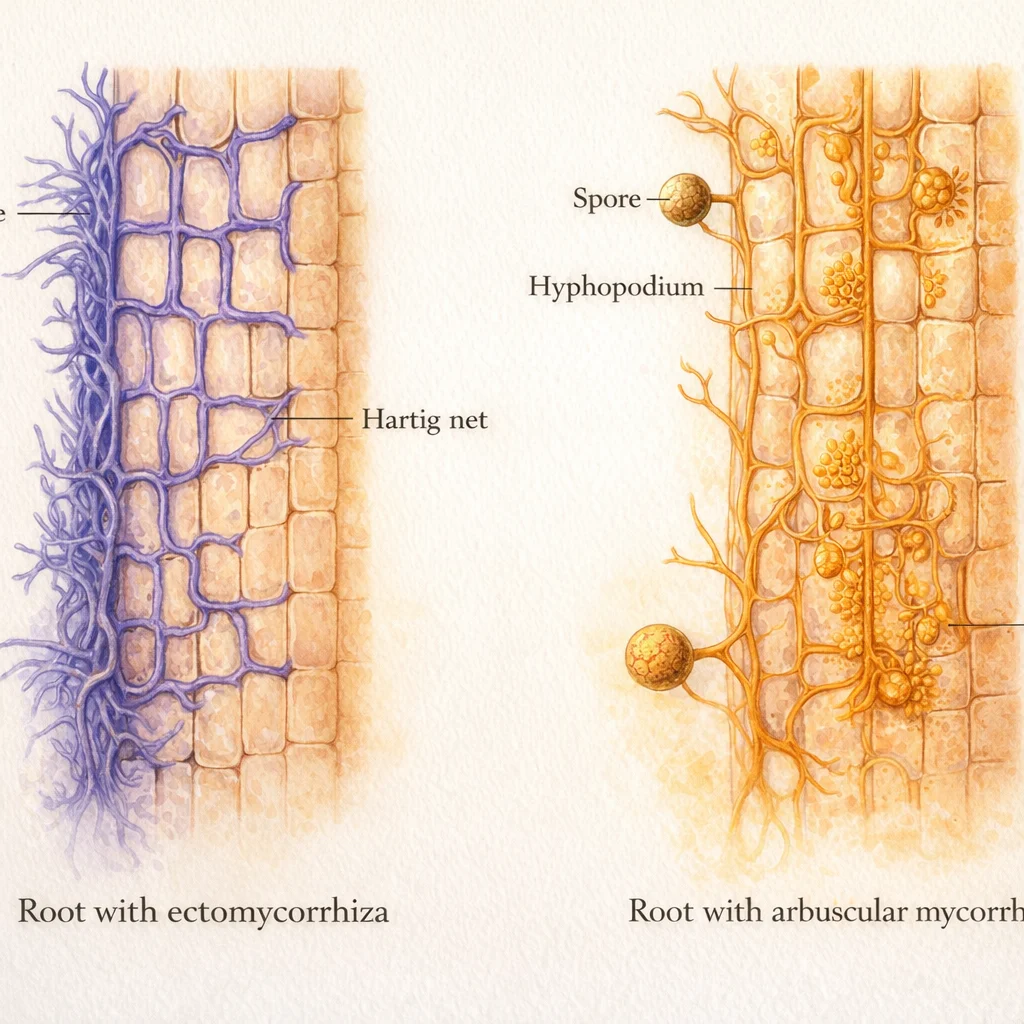
This partnership is older than trees. The earliest fossils of mycorrhizal roots date to 407 million years ago, placing them very early in land-plant history (Brundrett & Tedersoo 2018). Over 80% of land plants — more than 250,000 species — still form these associations today (Martin & van der Heijden 2024). They come in four distinct forms: arbuscular (AM), ectomycorrhizal (EcM), ericoid, and orchid mycorrhizas, each built from different fungi with different root architectures. Only about 10% of plants go it entirely alone.
What the fungi provide is remarkable in its quiet scale. Up to 90% of a plant's phosphorus and nitrogen can arrive through mycorrhizal hyphae rather than through the roots themselves (van der Heijden et al. 2008). In return, plants channel 5–30% of their photosynthate underground to sustain the fungal network — a flow so large that globally, the carbon entering mycorrhizal mycelium equals roughly 36% of annual fossil fuel CO₂ emissions (Hawkins et al. 2023). The hyphae also bind soil particles into aggregates, building the physical structure of the earth beneath your feet.

The Mycorrhizal Balance
Luo et al. (2023) analysed 74,563 forest inventory plots across 35 US ecoregions and found that in ~77% of ecoregions, forests mixing arbuscular mycorrhizal (AM) and ectomycorrhizal (EcM) host species were more productive than forests dominated by either type alone. The mixing benefit was most pronounced at low species richness (≤5 species) — the range most directly comparable to designed plant guilds.
In tropical gardens: most plants are AM hosts by default. To activate the mixing benefit you must deliberately include EcM-associating species — dipterocarps (meranti, shorea), tropical oaks, or Casuarina. Without at least one, the mycorrhizal mixing component contributes nothing to your score.
Mycorrhizal type assignments follow Brundrett & Tedersoo (2018), which compiled the family-level mycorrhizal classification used globally.
| Type | Common families | Soil layer | Nutrient form |
|---|---|---|---|
| AM | Most herbs, tropical trees, crops | Mineral | Soluble P, mineralised N |
| EcM | Pinaceae, Fagaceae, Betulaceae, Dipterocarpaceae | Organic | Organic N & P (enzymatic) |
| NM | Brassicaceae, Chenopodiaceae, Proteaceae | — | No mycorrhizal partner |
Mycoparasitic Disease Suppression
Trichoderma, Clonostachys, and related fungal species are the soil's own disease fighters — free-living fungi that hunt and dissolve soil-borne pathogens. Unlike mycorrhizal fungi, they don't form nutrient-exchange partnerships — but they can colonize root surfaces, trigger plant defences, and reduce the population of disease-causing fungi around your plants' roots. When Trichoderma strains are deliberately applied as inoculants, pathogen reductions of 5–87% have been documented depending on the strain, target pathogen, and host crop — with the strongest results in this review against Fusarium species and root-rot pathogens (Singh et al. 2024, a review of biocontrol studies).
Critically, Trichoderma discriminates between beneficial fungi and pathogens at a distance (Stange et al. 2024) — growing toward pathogens while avoiding ectomycorrhizal fungi in laboratory assays. A guild designed for rich mycorrhizal connections and one with active Trichoderma aren't in conflict — they coexist without interference.
Chemical detection at a distance
Trichoderma detects fungal pathogens at a distance — before any physical contact. It grows toward pathogens while showing the opposite response to ectomycorrhizal fungi. This targeted response is why it behaves more like a biocontrol agent than a blanket antifungal (Stange et al. 2024).
Attack and dissolve
Trichoderma secretes chitinases and glucanases — enzymes that break down the cell walls of pathogens like Fusarium, Rhizoctonia, and Pythium. Singh et al. (2024) documented up to 87% pathogen reduction, though efficacy varies widely by strain and target.
Spares mycorrhizal networks
In laboratory assays, Trichoderma actively grew away from EcM fungi and did not activate its mycoparasitic genes against them — reserving its dissolving enzymes for pathogens (Stange et al. 2024). This suggests disease suppression and mycorrhizal diversity can coexist, though field confirmation is still limited.
Nitrogen-Fixing Bacteria
Nitrogen is often a limiting nutrient in plant growth. Plants cannot use atmospheric nitrogen directly. Nitrogen-fixing bacteria solve this problem by breaking the N≡N triple bond and converting atmospheric nitrogen into ammonium (NH₄⁺) that plants can absorb.
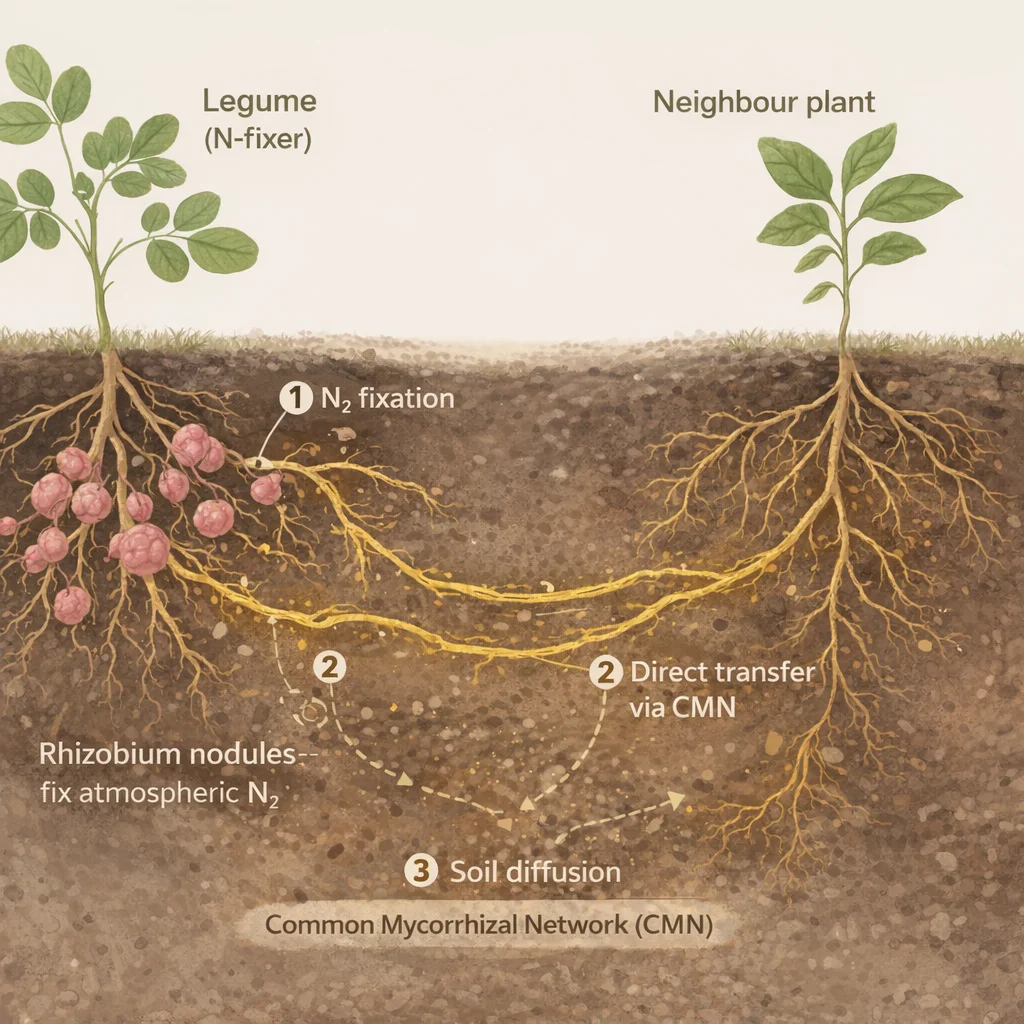
The most important partnership in a garden guild is between legumes (Fabaceae) and Rhizobium bacteria. The plant provides carbon-rich photosynthate to fuel the bacterium; the bacterium provides fixed nitrogen in return. Root nodules are the physical site of this exchange. Inside them, rhizobial bacteroids use nitrogenase to convert atmospheric N₂ into ammonia, which the plant then turns into ammonium (NH₄⁺). Leghemoglobin helps keep oxygen levels low enough for that reaction to work.
Legume–Rhizobium
The dominant pathway. Many Fabaceae (beans, peas, clover, acacia, soy) form nodules with specific Rhizobium strains, and the exact outcome depends on the host, the bacterial strain, and the soil environment.
Non-Legume Symbiosis
Frankia bacteria partner with alder (Alnus) and casuarinas. Less common than legume nodules, but still important in some woody lineages. Allocasuarina is unusual — it also forms EcM associations (Brundrett & Tedersoo 2018), giving it dual soil benefits.
Free-Living Fixers
This page's score focuses most strongly on the better-studied symbioses with legumes and actinorhizal hosts. Free-living nitrogen fixers are not the main driver of this metric.
Why legumes score so high: legumes can contribute nitrogen to the guild over time — shared through root turnover, nodule decomposition, and movement into the surrounding soil.
Infection and nodule formation
Rhizobium bacteria in the soil detect flavonoid signals released by legume roots. The bacteria enter through the root hairs, forming an infection thread that travels deep into the root where nodules develop.
Nitrogen fixation
Inside nodules, bacteroids express nitrogenase — the enzyme that breaks the N≡N bond. Leghemoglobin helps regulate oxygen levels so the anaerobic reaction can proceed. The product is NH₄⁺.
Nitrogen release to the guild
Fixed nitrogen enters the soil as roots leak it, as old nodules break down, and as fallen leaves decay. Neighbouring plants absorb this nitrogen directly — free fertiliser for every plant sharing the same soil volume.
How We Score It
Your soil biodiversity score combines three things: whether your guild hosts both types of mycorrhizal fungi, how many nitrogen-fixers it includes, and whether any plants are known hosts of disease-fighting fungi.
The scoring curve peaks at a 50:50 AM:EcM mix. The exact optimum isn't specified in the paper — we use 50:50 because both fungal types access different nutrients in different soil layers, and their benefits are largely complementary.
What We Can't Know Yet